Don Miller, Don Miller Land Use, Gisborne, New Zealand
Introduction
The Gisborne region has the worst soil erosion problems in New Zealand, mainly because of its soft rock geology. Vetiveria zizaniodes was introduced there in 1994 to test its usefulness for erosion control in a temperate climate.
Gisborne is an eastern coastal region at latitude 38 0 south and has cool winter (min air temp. -6 0 C ) and warm summer (max air temp. 38 0 C ) temperatures. The mean air temperature is 14.1 0 C and sunshine hours average 2180 hours per year. The average rainfall is 1060 mm but ranges from 1537 mm to 668 mm.
The first large scale plantings of Vetiver Grass in Gisborne were in an outdoor nursery (February 1994) and excellent growth was achieved in the first and second years. (Because the growing season was nearly finished in February the young plants were covered in clear plastic tunnels for the first 9 months) The plants reached about 1.6 metres in height and a diameter of approximately 200 mm by the end of the second growing season (March 1995).
Growth during the third summer (November 1995 to March 1996) was greatly reduced and by the end of the 4th growing season (March 1997) many of the original plants were dead or had only a few live tillers. A new nursery had been established by then so plant material for propagation was still available from the younger plants.
The author wishes to stress that the research described below was carried out to determine the cause of poor growth or death of vetiver in Gisborne and to find practical ways of overcoming the problem. It was not intended to be published as a scientific paper but is presented as information to assist other vetiver users.
1. Soil Temperature
During the initial introduction of Vetiver Grass to New Zealand both Richard Grimshaw and John Greenfield (who had developed the use of Vetiver in Fiji in the 1950's) discussed the effect of soil temperature on vetiver growth. They indicated that below about 22 0 C growth was slow and that below about 19 0 C growth ceased.
During the three growing seasons of early 1997 to early 1999 a number of short term studies of soil temperature were undertaken in the nursery area, to see if this was the major factor affecting vetiver grass survival.
Trial 1
In January 1997 temperature readings at 75 mm depth were as follows:
Under bare ground 23.5 0 C
Under 2 year old vetiver 19.0 0 C
Under 3 year old vetiver
with poor vigour 17.0 0 C
Trial 2
These results indicated a relationship between ground cover, soil temperature and vetiver health. In December 1997 areas of one year old and some areas of the now almost dead original vetiver plants were mowed to a height of about 60mm. On a part of the mown area all leaf litter was raked and removed to give bare ground.
In February 1998 at 7:00 pm after a day with air temperatures up to 33 degrees, soil temperatures at 50 mm depth under the three ground covers were:
Bare ground 32 0 C
Mown grass mulch 27 0 C
Un-mown vetiver 22.5 0 C
The recovery of the mown, raked vetiver of one year age was rapid and complete. The original vetiver plants, now almost 4 years old, gradually regrew where there were still green shoots.
Trial 3
The rate at which soil temperature rose after mowing and raking was shown on a clear day in late October 1998 (spring in the southern hemisphere) when soil mown and raked at 1:00 pm showed the following temperature rise by 4 pm:
1:00 pm 4:00 pm
30 mm depth 22 0 C 30 0 C
80 mm depth 18 0 C 24 0 C
Trial 4
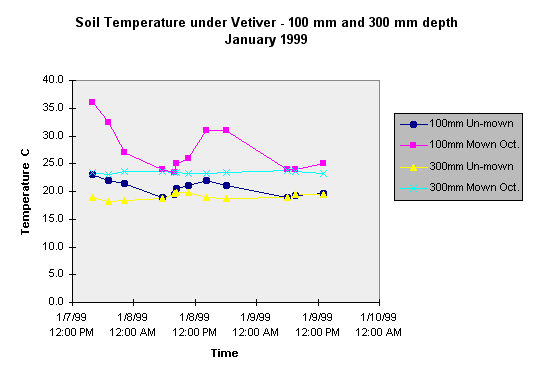
In early January 1999 thermistor units were used in conjunction with thermometers to obtain soil temperatures at depths of 100 mm and 300 mm under two year old vetiver mown and raked in October 1998 and under un-mown four year old vetiver in poor health.
The results for a two day period with air temperatures up to 36 0 C are summarised in the following graph.
This shows the large diurnal variation of temperature at 100 mm depth in both mown and un-mown sites while the fluctation at 300 mm depth is damped. At both depths the temperature is substantially higher under the mown and raked surface, despite the two month regrowth of vetiver leaf at that site.
Note that the apparently critical temperature of 23 0 C is exceeded at the 300 mm depth in the mown and raked plot for the two days.
Trial 5
To further investigate the rate of temperature change under different ground cover a brief study was carried out in October 1999. Two year old vetiver that had not been previously mown was given three treatments: One section was left un-mown, another mown with the leaf litter left on the ground and the third mown and raked clear.
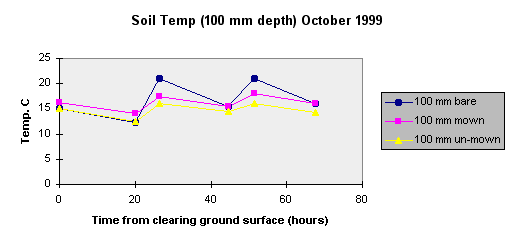
The layer of mown leaf litter reduced peak late afternoon soil temperatures but tended to reduce night time heat loss. The effect of bare ground on growth will depend on the ability of vetiver grass to respond to brief periods of high soil temperatures.
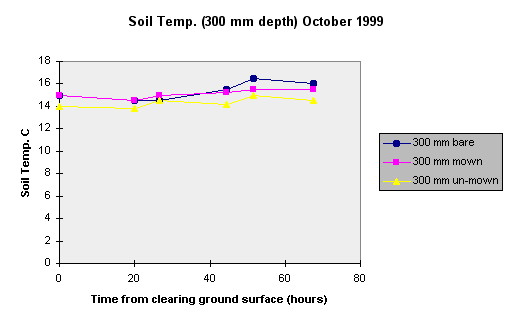
The temperature changes at 300 mm depth are less than at 100 mm but an increase of 1 0 C occurred in the mown and raked treatment over the 68 hour trial period. This could have a significant effect on root growth. The other two treatments had a rise of 0.5 0 C.
The leaf re-growth appears to be similar on the two mown treatments with leaf extension up to 180 mm in 23 days. Growth of the un-mown vetiver grass was not obvious.
Rooting Depth
The depth of vetiver roots and their strength is important in embankment stabilisation. If soil temperature at depth is too low for healthy root growth the increase in strength may be minimal.
In January 1994 the temperature in the base of an excavated hole 1.0 metre deep was 19.0 0 C. Vetiver roots from a 2 year old test planting were present at 700 mm depth at a density of about 400 per m2, although the diameter was very small. Two larger roots could be identified by their smell, implying that they were in their second year. At 1.0 m depth there were still significant numbers of roots found.
If the method of temperature measurement was accurate it indicates that vetiver roots will grow in soil at 19 0 C, although at a much lower density than observed in tropical soils.
In late September 1999 the 1.0 metre depth soil temperature (thermocouple probe) under 3 year old vetiver was 12.5 0 C. This would possibly be the low point on the annual temperature cycle at that depth.
Digging of plants to obtain tillers in the 3rd year revealed a number of cicada (Hemiptera : Cicadidae) nymphs in the soil around the roots. At that time they were not thought to be significant.
During the clearing of leaf litter in Trial 2 (December 1997) a layer of soil was accidentally removed and many cicada holes about 10 mm in diameter were revealed. Each hole appeared to contain a live mature cicada nymph which almost immediately emerged onto the ground surface to hatch.
There are about 30 species of cicada in New Zealand. From their colour and stage of development these appeared to be the most common species, Amphisalta zelandica. This insect lives in the soil as a sap sucking nymph for 1 to 5 years living off roots before it burrows to the surface and emerges around mid-December. It can tunnel as deep as 400 mm into the soil while it sucks xylem sap from roots.
The tunnels appeared to be connected to the surface by a small breathing hole about 20 mm long. A layer of soil about 20 mm thick was removed from the trial site to expose the remaining tunnels and they were counted. From counts of 9 plots of 0.09 m2 area a population estimate of 295 cicada tunnels per m2 was made. This seemed to be an extremely high number of cicadas. As there may be more than one nymph per tunnel the actual population may have been higher.
Within 20 metres of this site a sample taken under pasture grass had a count of up to 60 tunnels per m2. Positive identification of the tunnels under the long established grass was more difficult than in the soil dug 4 years earlier. Current research in New Zealand woodlands has found populations averaging about 20 cicadas per m2 (J. Perrott, 1999. Pers. com.).
An American study (Dybas and Davis, 1962) reported 300 cicadas per m2 as being the highest biomass reported for any naturally occurring terrestrial animal.
The population of 295 cicadas per m2 found in the vetiver nursery was therefore very high by world standards and was almost certain to be affecting the health of the vetiver grass plants by damaging their roots. Even stronger evidence was the obviously increased concentration of holes close to the vetiver plants themselves.
Discussion
It seems from these observations that both cicada attack and low soil temperature could be responsible for the early death of vetiver grass plants in Gisborne.
Soil temperature may not be the only explanation for the plant death. Several isolated vetiver grass plants near the nursery have survived for over 6 years now with slow steady growth. Soil temperatures there would not be any higher than those reported above but cicada have not been observed in those sites.
It is also seen in trial 5 that vetiver leaf growth is occurring already in early October 1999 with soil temperatures of 16 0 C. Leaf growth may, however, occur at lower soil temperature than root growth.
Cicadas are insects found in all tropical regions and if attack on vetiver grass was a common problem it would have been reported.
A more likely reason is that in the cool soil conditions that result from shading by the vetiver plants, root growth slows to a low level. The amount of sap being taken by the cicada may prevent any significant growth at all.
The roots of most grasses apparently live for about two years. The roots that grow in the early stages when the soil is heated by direct solar radiation would then sustain the vetiver plant for two years. It is surmised that by 2 years the cicada population may have increased after the initial low levels resulting from cultivation. The cicadas may be concentrated on the vetiver roots, possibly attracted by the smell. With the death of the 2 year old roots, the cooler soil conditions slowing replacement root growth, and the sucking of sap by cicadas from those new roots, the health of the vetiver plant quickly declines.
If this theory is correct mowing to allow solar radiation to heat the soil might only be needed once every two years, particularly if shallow cultivation was able to interrupt the life cycle of the cicada.
An additional factore may be that the close spacing of plants in the nursery might shade out new tillers shooting from the base of the vetiver. This has not been investigated.
Recommendations
Where vetiver is dying mysteriously in warm soil conditions it might be wise to check for the presence of high cicada populations. In the Japanese region of Isigaki-Zima the local cicada, Mogannia minuta, reached epidemic status in sugar cane (up to 350/m2 ) and stayed at high populations for at least 10 years (Yosiak Ito and Masaaki Nagamine, 1981). The population in near-by grassland remained at normal low levels with some migration from the sugar cane fields. That study concluded that escape from normal predator controls might have allowed the rise in cicada population. Once those high numbers were established they became self perpetuating.
It is possible that isolated high populations of cicadas can arise and persist for some time. Population assessment might involve upper soil removal and tunnel counting, observations of nymph exoskeletons on vetiver stems or the taking of soil core samples. Insect traps could also be used.
Where cool soil conditions are a problem and there is adequate solar radiation, vetiver growth may be enhanced by mowing at the start of the growing season. Vetiver grass mown to 50 to 100 mm height may not trap sediment efficiently and a higher cut with side trimming may be more effective. A higher cut would allow stem nodes to remain as potential new plants if burial with sediment occurs.
Higher soil temperatures can be obtained with leaf litter removal but the value of that would have to be determined for each situation.
The minimum winter temperature is often regarded as a major cause of vetiver death. In some cases, however, low summer maximum temperatures may be the main reason for poor growth and death.
Bibliography
Dybas, H. S., and D. D. Davis. 1962. A population census of seventeen-year periodic cicadas (Homoptera: Cicadidae: Magicicada) Ecology 43: 432-444
Perrott J., Massey University, Palmerston Nth, NZ
Yosiak Ito and Masaaki Nagamine. 1981. Why a cicada, Mogannia minuta Matsumura, became a pest of sugarcane: an hypothesis based on the theory of `escape'. Ecology 63 (2): 321-328