| Grass Barriers In Cassava Hillside Cultivation: Rooting Patterns And Root Growth Dynamics |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
K. Tscherning a, D.E. Leihner a,*, T.H. Huger a, K.M. Muller-Samann b, M.A. El Sharkawy b Institutfiir Pflanzenproduktion in den Tropen und Subtropen, Universitat Hohenheim, 70593 Stuttgart, Germany' b Centro Internacional de Agricultura Tropical (CIA T), Apartado Aereo 6713, Cali. Colombia Received 20 September 1994; accepted 23 May 1995. Field Crops Research 43 (1995)131-140 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abstract Living grass barriers can effectively reduce soil erosion on hillsides under cassava [Manihot esculenta Crantz]. Competition below ground in barrier crop systems is, however, poorly understood. The objectives of this study were, therefore, (i) to describe rooting patterns and spatial root distribution of cassava and of grasses commonly used as live barriers in soil conservation, and (ii) to determine root growth dynamics over time, using direct observation and quantitative methods. Field research was carried out near de Quilichao, Colombia, on a Typic Dystropept soil where average annual precipitation is 1799 mm. Single 20-m rows of vetiver grass Vetiveria zizanioides (L.) Nash, lemon grass [Cymbopogon citratus (DC. ex Nees) Stapf] and guatemala grass [Tripsacum andersonii J.R. Gray] were grown on a 13% slope. Cassava was planted in rows on either side of the grass barriers. Observation pits were dug in the cassava-grass plots and perspex sheets fitted closely to exposed walls. These pits allowed periodic tracing of roots. Root length was recorded in metres either per total of an exposed surface measuring 0.8 (depth) and 1.2 m (width) or measuring 0.2 (depth) x 1.2 m (width). Because of their vertical growth and their weak tendency to branch, the roots of vetiver grass rarely mixed with cassava roots, signifying a largely separate soil exploration by the two plants. In contrast, the profuse vertical and horizontal spread of guatemala grass and cassava roots indicated a joint exploration of soil volume. The cassava-lemon grass system revealed an intermediate pattern. The longest roots per exposed area of 0.2 x 1.2 m were observed for cassava growing on both sides of the vetiver grass barrier, and increasing from 6.0 m in the ~20 cm profile segment to 7.3 m in the 2040 cm segment. In contrast, roots of cassava with guatemala grass shortened from 5.4 m in the uppermost profile segment to 3.6 m in the 2040 cm segment. Vetiver grass exhibited the slowest and steadiest root growth and Guatemala grass the latest. Final total root lengths obtained from the entire exposed soil profile were 7 m for vetiver grass, 16 m for guatemala grass and 17 m for lemon grass. Cassava root growth was very slow initially, but increased 25-28 weeks after grass planting when grass root growth temporarily slowed down as a result of cutback. Final total cassava root lengths obtained from the exposed soil profile were 17, 10 and 10 m in association with the three grasses, respectively. The differences in rooting pattern, root growth dynamics and final root lengths suggest that different degrees of root interaction may have occurred. with possible consequences for soil exploration, and nutrient and water uptake. Key words: Cassava; Grasses; Competition; Grass barriers; Root growth; Rooting patterns 1. Introduction Cassava adapts to a variety of ecological conditions and tolerates low soil fertility. drought and pests, rarely experiencing complete crop failure. This is why the crop continues to hold an important position in traditional tropical cropping systems of small farmers, who are increasingly displaced to marginal, sloping lands that are unsuitable for large-scale mechanized cultivation. Cassava cultivation on slopes steeper than 15%, together with highly erosive rains, frequently leads to high soil losses. Burgos (1987) measured soil losses of 101-111 t ha in recently tilled cassava plots in Costa Rica and recommended intercropping cassava with fast-growing crops. Soil losses of the same magnitude were recorded by Howeler (1985) in cassava plots on slopes of 40 to 50% in the Central Cordillera of Colombia. In these trials, erosion was drastically reduced by either mulching with maize straw or interplanting strips of well-adapted grasses. Several studies discuss the benefits and problems of grass barriers for soil protection in long-season crops such as cassava and coffee (Lal, 1977; Webster and Wilson, 1980; Aristizabal, 1988; Cadavid-Lopez, 1990). None, however, quantitatively assess crop-barrier interactions, using growth patterns both above and below ground, across space and time. Knowledge of the underground structure of plant communities is necessary to understand their ecological interaction (Schubert, 1982). For barrier-crop interactions, this understanding is particularly significant for interpreting the ecological and agronomic success or failures of soil conservation technologies and their adoption or rejection by farmers. In this context, estimating possible degrees of below-ground interaction for individual cassava-grass combinations thus appeared important. The objectives of this study were, therefore, (i) to describe rooting patterns and spatial root distribution of cassava and grass species commonly used as live barriers in soil conservation, and (ii) to determine root growth dynamics over time by quantifying weekly and cumulative root length increases. 2. Material and methods 2.1.Experimental conditions Field research was conducted near Santander de Quilichao, Colombia, at the research farm of the Centro Internacional de Agricultura Tropical (CIAT). The area has an average annual temperature of 23.8�C. Rainfall and evaporation data for 1972-1988 indicate that the average annual rainfall of 1799 mm exceeds the annual potential evaporation of 1589 mm. However, during June through September and in January, potential evaporation exceeds precipitation. Rainfall intensity occassionally exceeds 100 mm h-1 and rainy periods can be interrupted by dry spells lasting a week or longer, even during peak rainy seasons. The experiment was carried out on a Typic Dystropept soil (IGAC, 1976), which has a clay texture, high organic matter, and low fertility (Table I). The field was located on a slope of 13%. Table1. Soil chemical properties of root-observation profiles for cassava-grass systems, Santander de Qailichno, Colombia
[
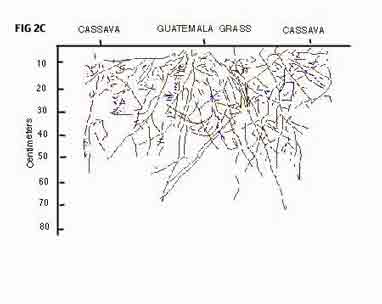
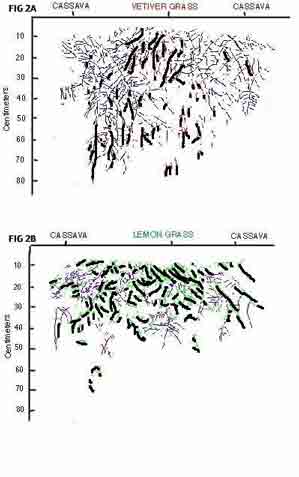
Fig. I. Planting arrangement of grasses and cassava in individual plots for root length and growth studies, 2.2. Grass and cassava planting On May 30, 1991, guatemala grass [Tripsocurn andersoni J.R.Gray], lemon grass [Cymbopogon citratus (DC. ex Nees) Stapf] and vetiver grass [Vetiteria zizanioides (L.) Nash] were planted in rows across the slope. Each grass species was planted in a row 20 m long, and separated from others by 1.9 m. Within each row, the tillers were planted at 10-cm intervals. After full establishment was reached on August 12, 1991, 10 weeks later, the soil between grass barriers vxaS loosened with a hoe and tiled with a rot~vator to a depth of 25 cm. Once the soil was prepared, grass was removed from the row every alternate 5 m. The cassava variety M Col 1505 was then planted in double rows between grass barriers at a density of IS 038 plants ha~ 'Untreated stakes, 20 cm long, were planted vertically at 55 cm from the grass row, leaving 80 cm between the two cassava rows. Along the row, the stakes were planted at 70-cm intervals. Thus, an individual plot (4.9 x 1.9 m, Fig. 1) consisted of 4.9 m of grass row (present or removed) with 7 cassava plants on either side of the grass barrier. Santander de Quilichan. Colombia, 1992. NOT INCLUDED] Each treatment was replicated twice, so the field design comprised 12 plots, half consisting of grass-cassava systems, half consisting of cassava sole crops established by removing the grasses at cassava planting time. No mineral fertilizer was applied, but the trial was irrigated during dry spells: 50 mm on August 16, 30 mm on August 28 and 30 mm on January 8, using a sprinkler irrigation system. 2.3. Grass barrier management The grass barriers were cut back by machete on November 18, 1991, 24 weeks after grass planting (WAGP); Guatemala grass to ground level, vetiver grass and lemon grass to 25cm. On April 1, 1992 (44 WAGP), guatamala grass was cut to ground level a second time. Vetiver grass and lemon grass were not cut to ground level, because they recover too slowly, especially in dry periods. 2.4. Root observations For two weeks after cassava planting, root observation pits - 1 m deep, 1.3 m wide and 1 m long - were dug in the "cassava with grass" plots only. One long wall of each pit spanned two rows of cassava with one grass row in-between (Fig. 1). This wall was made vertical with a metal blade specifically made for this purpose. Small unevenesses in the profile wall were corrected with a spatula before a 5-mm-thick acrylic sheet, measuring 1.27 x 1.00 m, was installed against it. When the wall was not in use, it was covered by a black plastic film to prevent light from affecting those roots growing against the acrylic sheet. In addition, a 60-cm-thick styrofoam board, measuring 2 x 3 m covered the whole pit to protect against fluctuations of light and temperature. A corrugated plastic sheet protected the pit from rain. Transparent 1.0 x 1.2 m acetate films were fixed to the acrylic sheets and, from September 3 (14 WAGP) to January 22 (34 WAGP), rooting patterns and root length increases were traced on the sheets at weekly intervals. Drawings were done with different coloured markers to distinguish roots of the two plant species and individual root growth increments from one week to the next. After tracing, the acetate films were taken to the laboratory, where the drawings were copied on semi-transparent paper, using various blue tones for cassava roots and various red tones for grass roots. Thus, by the end of the observation period, a complete picture of both cassava and grass rooting patterns was available. The last drawing showing cumulative root length, was used to obtain information on root length distribution. A transparent acetate film, marked with a 15 x 20 cm grid, was placed on top of the drawing and root lengths were measured in each grid, using a hand-held distance scanner (Fa. Thoma, Erlangen, Germany). The grid was placed so the centre of the grass barrier could be taken as the zero reference point. Starting from zero, root lengths were measured every 15-cm segment up to 60 cm on both sides of the grass barriers and every 20 cm in depth down to 80 cm. Hence, root length was recorded in metres either at a depth of 20 cm across a width of 1.2 m (or 15-cm portions thereof), or within the total exposed surface of 80cm (depth) x 1.2 m (width). The fine roots of cassava and grasses were easily distinguished according to (i) the ramification pattern, in which cassava lateral roots typically start at right angles from the major roots, whereas grass lateral roots begin at narrow angles, (ii) vetiver grass roots are light cream and lemon grass roots light yellow, whereas cassava roots are intensely white, and (iii) grass roots, particularly those of vetiver and guatemala grass, were considerably thicker than cassava roots. 2.5. Data analysis To reproduce rooting patterns, drawings derived from replicate 2 of each treatment were used because no obvious differences between drawings from replicate 1 and replicate 2 could be observed. The analysis of variance for root distribution was calculated with the General Linear Models (GLM) procedure of SAS (1988). The lowest depth (60-80 cm) was not included in the analyses because most observations were zero. In the model, grasses were treated as a random variable, and soil depths and distances as repeated measurements. To avoid a third repeated-measures factor, the values from both sides of the grass barrier were averaged before calculating the analysis of variance. 3. Results 3.1. Rooting patterns Distinct root development and distribution patterns occurred for each cassava-grass combination. Both replicates produced very similar rooting patterns. The least intermixing of the two root systems was found in the cassava-vetiver grass combination (Fig. 2A) While cassava roots spread profusely in both lateral and vertical directions, roots of vetiver grass showed clear tendency to grow vertically down the profile witl little lateral spread or tendency to branch. However where cassava and vetiver roots met, they grew close together over long distances with no signs of root avoidance. A good downward root penetration of both species was found over the whole depth of the profile. In the cassava-lemon grass plots, both species showed profuse root development only in the upper part of the profile (Fig. 2B). In the lower part of the profile, there were few cassava or lemon grass roots. The dense growth and ramification of lemon grass root apparently did not prevent cassava roots from also exploring the upper segment of the profile. However in the grass area (centre of figure) cassava roots were noticeably less dense than in the cassava areas (left and right side of the figure) and, equally, grass roots were fewer under the cassava rows than under the grass row. The rooting pattern of the cassava-guatemaIa grass combination was different again, showing strong penetration of grass roots in all directions of the profile (Fig. 2C). Cassava roots developed their usual strong lateral and vertical spread, and close contact was observed between cassava and grass roots. The root system of guatemala grass was vigorous, deeply penetrating and consisted of many thick roots with little ramification.3.2. Ro lengths and distribution of grasses
Root growth of all three grasses was greatest in the 0-20 cm segment of the exposed profile, with few differences between upslope and downslope root development (Fig. 3 NOT INCLUDED). By the experiment's end, root lengths were 2.70 m for vetiver grass. 7.49 m for guatemala grass and 9.04 m for lemon grass. Vetiver gras typically produced 96% of its root length to a distance of only 30 cm on each side of the barrier. In contrast the root lengths of the two other grasses decreased only gradually with increasing distance from the grass barrier. At 20-40 cm depth, root lengths were shorter than those in the first 20 cm of the profile: lengths were 1.64 m for vetiver grass, 5.65 m for guatemala grass, and 5.88 m for lemon grass. Both guatemala grass and lemon grass roots were longer on the upslope of the grass barrier than on the downslope. In the 40-60 cm segment, root length was further diminished, with the strongest reduction in lemon grass (88%) (Fig. 3). Very few lemon grass and guatemala grass roots could be found in the lowest segment (60-80 cm), and root lengths were only 15 and 39 cm, respectively. Consistent with its charactenstic rooting pattern, vetiver grass showed considerable root growth at this depth, with a total root length of 1.37 m, most of which was dowoslope. Statistical analysis of grass root length distribution showed a significant (P<0.05) "depth x grass species" interaction, confirming that the grasses have dif-ferent exploration pattems with depth. 3.3. Root length. ond distribution of cassava Cassava tended to produce more roots away from the grasses than near them (Fig. 4). In the 0-20 cm segment, the longest cassava roots were found in the cassava-vetiver grass combination, totalling 5.99 in. followed bv cassava lemon grass with 5.41 in and cassava guatemala grass with 2.83m. (fig 4). in the 20-40 cm segment, root lengths of cassava with lemon grass and guatemala grass were somewhat reduced, whereas roots ofcassava with vetiver grass were longer, totalling 7.31 m - more than 1.3 m longer than in the top 20 cm. The increase occurred mostly downslope. In the 40-60 cm segment, cassava root growth was further reduced but, again, the total root length with vetiver grass (3.36 m and 76% down slope) was considerably greater than in cassava-guatemala grass or cassava-lemon grass (2.00 m and 0.71 m respectively) (Fig. 4). Few cassava roots reached the 60-80 cm segment, except for those of cassasa with vetiver grass, where the downslope length was 86 cm. Cassava root lengths were statistically significant (P<0.0l ) for the ''distance x grass species'' interacation, indicating that the type of grass significantly influences lateral exploration by cassava roots. 3.4. Root grwoth dynamics Cumulative grass root lengths over time (Fig. 5) showed further differences in root growth dynamics, vetiver grass roots grew slowly and steadily, reaching 7 m for the total exposed surface, the smallest of the three grasses. Both lemon and quatemal grass roots grew more rapidly during the period 16-24 WAGP. Lemon grass roots started earlier, resulting in significantly greater root lengths, compared with vetiver grass at 17 and 19 WAGP. Guatemala grass, however, was slower, reaching similar values at 20 WAGP, followed by almost no growth at 25-29 WAGP, possibly because of the first cutback. After the cutback, both lemon grass and guatemala grass resumed rapid root growth. The total final root length of each of these two grasses was as much as two and a half times that of vetiver grass. During the active early growth of grass roots, cassava roots grew very little, becoming active only during the lag phase of grass root growth after cutback. With vetiver grass, cassava roots began growing rapidly as much as 3 weeks earlier than with the other two grasses (data not shown), resulting in a total final root length of 17 m, which is 70% greater than that in either of the other two cassava-grass combinations. 4. Discussion 4.1. Soil penetrability The soil was readily penetrated by roots down the whole depth of the observed profiles. Roots did not encounter physical obstacles, such as a hard pan or clay layer, that inhibited root growth or deflected roots laterally. Neither were there chemical barriers, root behavior being consistent with increasing pH and decreasing aluminum availability at lower depths. A concentration of plant nutrients in the first 40 cm probably explains why most grass and cassava roots were found exploring only the upper part of the profile. 4.2. Rooting pattern The greatest spatial separation of grass and cassava roots was observed in the cassava-vetiver grass combination. This resulted from the predominantly, downward direction of root growth in vetiver grass which occupied a root zone resembling an inverted ''U" centred below the grass row. A near-complete separation of grass and cassava roots was observed in the top 40 cm of the profile. This was associated with cassava's greatest root length, particularly at 20-40 cm. Further-more, cassava roots spread both laterally and vertically throughout most of the profile, suggesting that vetiver grass roots did not inhibit cassava roots from exploring the same soil volume. The root-zone separation in the cassava-lemon grass combination was not so clear. Lemon grass showed more lateral expansion than did vetiver grass. Cassava roots also spread laterally but root density in the area below the grass row decreased. The lower part or the profile was less intensely explored by lemon grass roots than by grass roots in the cassava-vetiver grass or the cassava-guatemala grass combinations. Cassava root penetration was also less deep, suggesting that unidentified minimal changes in soil chemical properties may have caused the shallow rooting. More so than in the other two combinations, guatemala grass roots aggresively expanded in all directions down the whole depth of the observed profile. Cassava roots also explored the whole profile, including the area immediately below the grass row. However, final total cassava root lengths were only about half of those observed in the two other combinations. In plant systems with strong and weak competitors, root systems of the weaker contponents are often reduced and their development is patticulatly inhibited in zones close to the stronger competitors (Baldwin and Tinker, 1972). This may be related to the depletion of water and nutrients around the roots of the more competitive species, which are then avoided by the root system of the weaker competitor. If no alternative soil volume can be explored, then normal root growth may be inhibited (Duncan and Ohlrogge, 1958; Klute and Peters, 1969; Rogers and Head, 1969; Trenbath, 1976). In our study, cassava roots did not avoid the soil volume explored by guatemala grass, but root growth was clearly reduced. This suggests that cassava is the weaker competitor in the cassava-guatemala grass system. 4.3. Root growth dynamics When comparing the dynamics of root growth of grass with that of cassava, periods of greatest activity were different. In the cassava-guatemala grass system, grass root growth was very active until 24 WAGP, contrasting with hardly any activity in cassava. This was followed by a period of less root growth in guatemala grass and more in cassava. Contrary to this pattern, vetiver grass showed a slow, steady root growth throughout the 14-34 WAGP period. This apparently allowed cassava roots to develop actively, starting as early as 25 WAGP, whereas in the other two systems, cassava roots showed an accelerated growth only after 28 WAGP. 5. Conclusions Based on these observations, we suggest that vetiver grass has the least competitive root system, with the shortest root length in the upper 40 cm of the profile and a root distribution that concentrates closely to each side of the grass barrier. These factors, together with a slow but continuous root growth over time, may have allowed cassava roots the most undisturbed exploration of the remaining soil volume, so producing the greatest total root length of all three systems. Both lemon grass and guatemala grass produced longer roots in the top 40 cm of the profile and showed a more active early root development up to 24 WAGP. However, only with guatemala grass did this lead to restricted cassava root growth, particularly noticeable in the top 20 cm. This spatial and temporal root growth pattern may indicate possible competitive effects as cassava is most likely to absorb most of its water and nutrients from this part of the profile. Despite the dense rooting of lemon grass in the first 20 cm, cassava root growth was scarcely reduced in this system, compared with the cassava-vetiver grass combination. This may have resulted from a synergistic effect that allowed cassava roots to explore, to some extent, those parts of the soil profile immediately beneath the grass barrier. Hence, both vetiver grass and lemon grass do not compete strongly with cassava, although for different reasons: the former for a non-interfering spatial distribution of roots and the latter for a possible synergistic effect. The guatemala grass thus remains the most aggressive of the components tested, possibly reducing the soil volume available for exploration and therefore of nutrients and water availability to cassava. References [Not included] |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||